Flantua S.G.A., O’Dea A., Onstein R.E., Giraldo C., Hooghiemstra H. 2019. The flickering connectivity system of the north Andean páramos. Journal of Biogeography 46:1808– 1825. 10.1111/jbi.13607
Please join our session and North American Paleontological Convention, held in UC Riverside this year, to explore how small fossils can make big contributions to our understanding of the past.
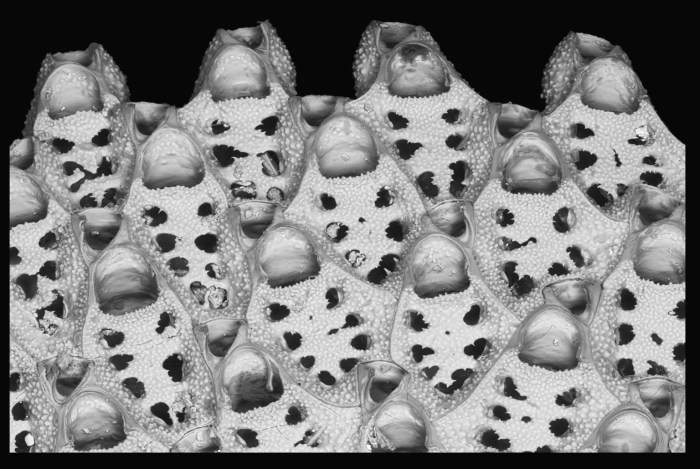
Close-up of the millimetre sized individual zooids of a colony of Discoporella
A quick look at the fossil record shows that no species lasts forever. On average, most species exist for around a million years, although some persist for much longer. A new study published in Scientific Reportsfrom paleontologists at the Smithsonian Tropical Research Institute in Panama shows that young species can take advantage of new opportunities more easily than older species: a hint that perhaps older species are bound to an established way of life.
“We’re lucky to live and work in Panama where nature has set up its own evolutionary experiment,” said Aaron O’Dea, STRI paleontologist. “When the Caribbean Sea was isolated from the Pacific Ocean by the slow uplift of the Isthmus of Panama, nutrient levels fell and Caribbean coral reefs proliferated. We can use the excellent fossil record to observe how Caribbean life responded to this environmental and ecological transformation.”
The team’s best choice for tracking the change was a peculiar family of marine animals known as the cupuladriid bryozoans. These relatively small animals consist of unusual, free-living, disc-shaped colonies of individuals called zooids. “Colonies form through sexual reproduction or asexually by cloning, as bits of the colony break off and continue to grow,” said STRI post-doc and coauthor Blanca Figuerola. “They abound on the sea floor along the continental shelf across the tropics, filtering plankton from the water via a beautiful waving crown of tentacles. When colonies die, their hard skeletons remain, and are exceptionally abundant as fossils.”
O’Dea’s group collected and identified more than 90,000 cupuladriid colonies from 200 fossil samples and 90 more recent samples collected by dredging the sea floor. The samples contained mud, sand, coral remains and other indicators of the kind of habitats where the bryozoans had lived. The team measured the abundances of the 10 most common species along gradients of these environmental and ecological indicators.
“We were intrigued to find that, even though all species could expand into the new Caribbean habitats created after final formation of the Isthmus, different species did so at different speeds,” said O’Dea. “The patterns were clear—old species that originated before 8 million years ago took 2 million years longer to expand into the new habitats than the younger species.”
“Perhaps younger species, which have smaller populations, are less tied to their history,” said former STRI post-doc and University of Saskatchewan researcher Santosh Jagadeeshan, another co-author. “Old species, with large, settled populations may be less able to escape from established roles and defined environmental tolerances because they mate with each other creating a high gene flow that makes it hard for genes for new traits to become established. It seems you can’t teach an old dog new tricks in evolution, either.”
The study was funded by Panama’s National Bureau of Science, Technology and Innovation, SENACYT, Panama’s National System of Researchers (SNI), the U.S. National Science Foundation (NSF), the Smithsonian Institution, STRI, the National Geographic Society and Mr. Josh Bilyk.
O’Dea, A., De Gracia, B., Figuerola, B. and Jagadeeshan, S. 2018. Young species of bryozoans occupied new Caribbean habitats faster than old species. Scientific Reports, DOI: 10.1038/s41598-018-30670-9
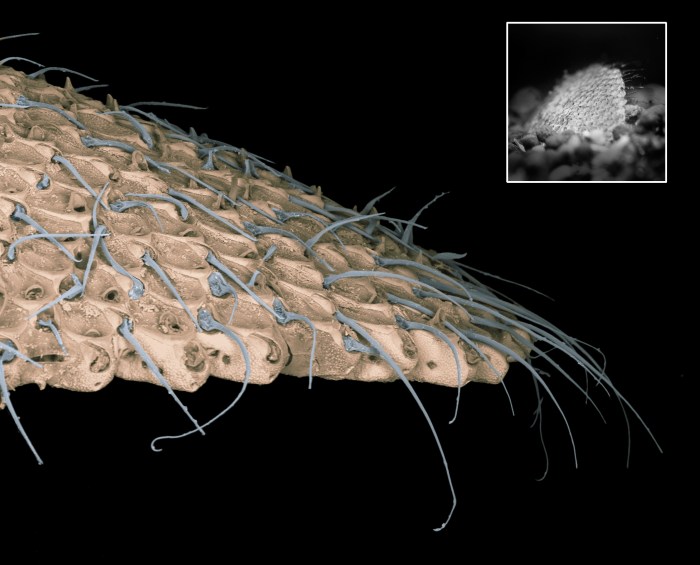
Stiff setae extend away from the edge of cupuladriid bryozoan colonies, and work in synchrony to allow the colony to “walk” over the sea floor and emerge from the sediment when buried
We thank Erkens and Hoorn for their constructive comments. Like us, they believe that collaboration between biologists, geologists and paleontologists focusing on data and analyses is required to unravel the history of the Isthmus of Panama. We agree with Erkens and Hoorn that the Continue reading “Building Bridges”→
The formation of the Isthmus of Panama stands as one of the greatest natural events of the Cenozoic, driving profound biotic transformations on land and in the oceans. Some recent studies suggest that the Isthmus formed many millions of years earlier than the widely recognized age of approximately 3 million years ago (Ma), a result that if true would revolutionize our understanding of environmental, ecological, and evolutionary change across the Americas. To bring clarity to the question of when the Isthmus of Panama formed, we provide an exhaustive review and reanalysis of geological, paleontological, and molecular records. These independent lines of evidence converge upon a cohesive narrative of gradually emerging land and constricting seaways, with formation of the Isthmus of Panama sensu stricto around 2.8 Ma. The evidence used to support an older isthmus is inconclusive, and we caution against the uncritical acceptance of an isthmus before the Pliocene.
From colleague and friend Paul Harnik’s Paleolab Blog: “How does environmental change shape the relationships between ecological traits and extinction risk? The fossil record is an invaluable resource for answering such questions. In a paper now available early online in the journal Global Change Biology, my collaborators and I show that over the last 500 million years global environmental and geochemical changes have had remarkably little effect on the relationship between geographic range size and extinction risk among marine mollusks. In other words, clams and snails with small geographic ranges have been at elevated risk of extinction throughout their evolutionary history regardless of broad-scale environmental conditions. In contrast, we found that mollusks that live in (rather than on) sediments on the seafloor tended to be at lower risk during times of warmer climate.”
We edited a special edition of the Bulletin of Marine Science entitled “Environmental, ecological, and evolutionary change in seas across the Isthmus of Panama”. This volume evolved from a colloquium Continue reading “Special edition on the Isthmus of Panama”→
An Isthmus is a narrow land bridge that joins two larger bodies of land. The world’s most famous isthmus, now split by the Panama Canal, connects the continents of North and South America and cuts the once continuous Tropical American seaway into two.
Upwelling along the Pacific coast of Central America today occurs where the land drops to less than 500m. Low land allows the trade winds to blow across and push surface waters out to open ocean causing strong coastal upwelling.
Records of seawater chemistry help constrain temporal variations in geochemical processes that impact the global carbon cycle and climate through Earth’s history. Here we reconstruct Cenozoic seawater Sr/Ca using fossil Conus and turritellid gastropods.
Our favored seawater Sr/Ca scenarios point to a significant increase in the proportion of aragonite versus calcite deposition in shelf sediments from the Middle Miocene, coincident with the proliferation of coral reefs. We propose that this occurred at least 10 million years after the seawater Mg/Ca threshold was passed, and was instead aided by declining levels of atmospheric carbon dioxide.
Pdf of the paper available by clicking on these images of cone shells…
Fossils and genes are the two principal ways to study evolution, but they are rarely studied together. This project allowed us to make the first integration of fossil and molecular records of cupuladriid bryozoans Continue reading “Integrating fossils and molecules”→
As a PhD student I devised and developed a completely new technique for investigating paleoseasonality. Reconstructions of paleoenvironments often fail to understand the importance of the mean annual range of temperature (MART) in both oceanographic and biological contexts. The new technique, called the ‘zooid size approach’ makes use of the temperature-size rule in colonial bryozoans to estimate MART. The temperature-size rule is a universal phenomenon that states that body size decreases as temperature increases.
At the time, our understanding of the temperature-size rule was rudimentary and it was necessary to develop hypotheses on the mechanisms behind the rule and then test them under controlled culture and natural experiments, before finally applying the approach to fossil bryozoans to estimate MART’s in ancient seas.
The original paper published in 2000 presenting the technique can be downloaded here.
Now 10 years later with my ex-Phd supervisor Beth Okamura we review the approach along with the growing body of work that has since been published on the theme. We consider the general issue of why body size varies with temperature, explore the limitations of the approach and highlight its advantages relative to other proxies for palaeotemperature inferences.
Download the pdf of this new paper by clicking on the image.
Even genetically identical animals can look very different if they grow and live in different environments. Think ‘you are what you eat’. I make use of this phenomenon to try to reveal changes in environments in the deep past by first understanding what drives change in morphology in the animals in question and then measuring that morphology in fossils through time.
I applied this paradigm to one of the most studied and certainly most discussed events in the history of life on earth. The K-T (Cretaceous-Tertiary) boundary, 65 million years ago and the demise of the non-avian Dinosaurs and a suite of other animals and plants in the seas and on land. I made detailed measures of morphology in a number of fossil bryozoans in a beautiful K-T section of chalk in Denmark.
Rapid and repeated changes in morphology suggest that there were a suite of environmental changes in the last few thousand years just before the K-T boundary.
Although we dont explore the causes of the extinctions, or the ‘smoking gun’, these results are important for a full understanding of the complex changes associated with major extinctions observed to occur around the world. Click on the image for the pdf.
A full understanding of any climate requires an appreciation of the amount of seasonal variation in temperature. This is important not only for present-day climatology but also for investigation of ancient environments. In this paper I present a novel approach to reveal how seasonal an ancient sea was by measuring the amount of variation in zooid size within colonies of fossil cheilostome bryozoans.



 Lauren Graniero, student at Texas A&M and STRI short term Fellow, just published another paper that helps us make sense of the significance of stable isotope ratios in skeletal material.
Lauren Graniero, student at Texas A&M and STRI short term Fellow, just published another paper that helps us make sense of the significance of stable isotope ratios in skeletal material. 





















You must be logged in to post a comment.