
Even genetically identical animals can look very different if they grow and live in different environments. Think ‘you are what you eat’. I make use of this phenomenon to try to reveal changes in environments in the deep past by first understanding what drives change in morphology in the animals in question and then measuring that morphology in fossils through time.
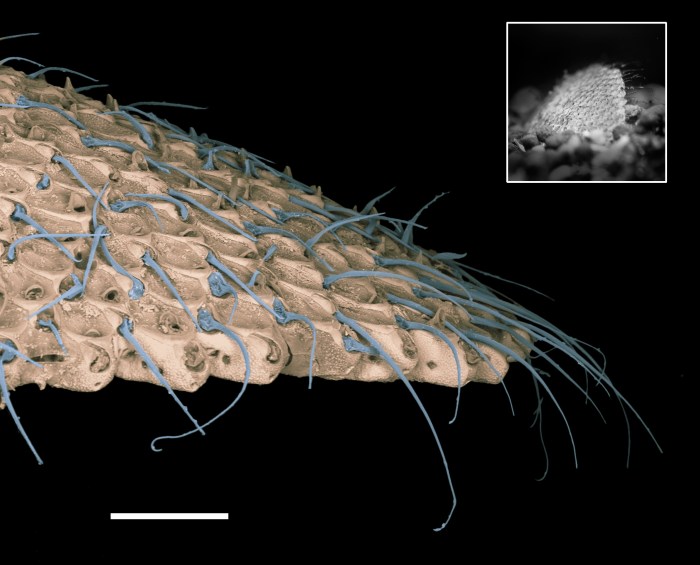
I applied this paradigm to one of the most studied and certainly most discussed events in the history of life on earth. The K-T (Cretaceous-Tertiary) boundary, 65 million years ago and the demise of the non-avian Dinosaurs and a suite of other animals and plants in the seas and on land. I made detailed measures of morphology in a number of fossil bryozoans in a beautiful K-T section of chalk in Denmark.
Rapid and repeated changes in morphology suggest that there were a suite of environmental changes in the last few thousand years just before the K-T boundary.
Although we dont explore the causes of the extinctions, or the ‘smoking gun’, these results are important for a full understanding of the complex changes associated with major extinctions observed to occur around the world. Click on the image for the pdf.














You must be logged in to post a comment.